Self-Explanation Strategies Enhance the Inhibition of Intuitive Misconception: Insights from ERP
published: 15 July 2025 | https://doi.org/10.63174/xdi.OETN9214
Abstract
To investigate the persistence of deeply entrenched intuitive misconceptions in science learning, this study examined the effectiveness and neural underpinnings of the self-explanation strategy (SES) in correcting the misconception that “ionic compounds consist exclusively of metal cations and nonmetal anions.” Participants were required to judge whether compounds qualified as ionic. By comparing behavioral and event-related potential (ERP) data between the Traditional Instruction Group (TIG) and the Self-explanation Strategy Group (SSG) when processing intuition-consistent (C) and intuition-inconsistent (IC) ionic compound stimuli, we found that in the TIG, C stimuli elicited larger P2 amplitudes, while IC stimuli evoked stronger N2 and LPP components. In contrast, the SSG exhibited reduced P2 amplitudes and enhanced N2 amplitudes under C conditions. These results indicate that persistent misconceptions require inhibitory control, and that self-explanation weakens early intuitive activation while enhancing conflict monitoring to promote inhibition.
1. Introduction
In the field of science education, researchers have long focused on the prevalent phenomenon of misconceptions among learners. Extensive studies demonstrate that students’ intuitive conceptions formed through daily observations prior to formal science education often exhibit systematic deviations from scientific knowledge[1]. For instance, students may hold deeply rooted intuitive beliefs such as “heavier objects fall faster” or “a single wire can light a bulb” [2,3]. Recent research has revealed that such misconceptions do not dissipate through scientific learning but rather coexist with scientific concepts[2,4,5,6,7]. This persistent coexistence can be explained through the dual-system theoretical framework, where the automatic processing of the intuitive system (System 1) competes with the controlled processing of the rational system (System 2) during scientific reasoning[8,9]. Neuroimaging studies have identified the crucial role of inhibitory control in attenuating interference effects from misconceptions[10,11,12]. This inhibitory mechanism substantiates the core proposition of the conceptual coexistence theory—that the essence of conceptual change in science education lies in establishing inhibitory dominance over misconceptions rather than conceptual replacement[13].
Within this context, generative learning strategies have gained prominence—the self-explanation strategy (SES) enables learners to bridge knowledge gaps, integrate prior and novel information, and trigger conceptual restructuring through cognitive conflict detection[14]. Notably, cognitive conflict itself has been neuro-scientifically validated as a pivotal cognitive event: When individuals experience such conflicts, it elicits inhibitory functions in relevant brain regions[15], ultimately establishing cognitive dominance of scientific representations. However, although existing behavioral studies have demonstrated SES facilitative effects on conceptual change, the precise mechanisms through which this strategy induces specific neural activity via cognitive conflict generation remain insufficiently explored.
Current research on conceptual change demonstrates a multimodal integration trend, combining behavioral assessments, neuroimaging techniques, and pedagogical interventions. Event-related potentials (ERPs), as an electrophysiological recording technique, offer superior temporal resolution and non-interference with task execution[16]. Recent years have witnessed ERPs providing novel perspectives for elucidating the neural mechanisms underlying conceptual change. For instance, Zhu et al.[5] found through an ERP study that increased P2 amplitudes to misconception-congruent stimuli, larger N2 amplitudes to misconception-incongruent stimuli, and enhanced late positive potentials (LPP) to stimuli conflicting misconceptions with scientific knowledge. These findings suggest intuitive misconceptions may persist after acquiring scientific knowledge. Similarly, Skelling-Desmeules et al.[6] employed ERP technology to demonstrate that adults’ judgments about “non-moving organisms” elicited enhanced prefrontal N2 and LPP components, reflecting persistent inhibitory demands against the “movement implies life” heuristic. These collective findings establish neurocognitive techniques as effective solutions for uncovering the cognitive neural mechanisms of conceptual change.
This study employs behavioral measures and ERP technology to investigate the persistence of intuitive conceptions regarding metallic compound composition in students’ cognition after acquiring scientific knowledge, while examining the intervention efficacy and neural mechanisms of the SES in misconception correction. In the experimental design, participants were allocated to either the Traditional Instruction Group (TIG) or the Self-Explanation Strategy Group (SSG), with continuous EEG recordings obtained during their validity judgments of presented stimuli.
Based on the misconception that ionic compounds consist exclusively of metallic and nonmetallic elements, ionic compounds can be classified into two categories: Type C (Congruent Stimuli) compounds composed solely of metallic and nonmetallic elements—which are correct according to both scientific concepts and the prior misconception—and Type IC (Incongruent Stimuli) compounds that do not strictly follow the simplistic combination rule of metallic and nonmetallic elements—which are scientifically correct but inconsistent with the prior misconception (Table 1). We hypothesize that persistent misconceptions exist in the brain. During the initial stage of scientific reasoning, the brain instinctively recruits these persistent misconceptions to assess stimulus plausibility: congruent stimuli match implicit memory traces, eliciting enhanced P2 components[5,17]; incongruent stimuli, however, elicit enhanced N2 components due to mismatch detection[18]. Subsequently, explicit retrieval of scientific knowledge is engaged to detect and inhibit this conflict. During this phase, incongruent stimuli involving conflict between misconceptions and scientific knowledge elicit increased LPP components [5,19].
Table 1. The Logic of Stimuli Sorting and Expected ERP Variations
| Congruent Stimuli (C) | Incongruent Stimuli (IC) | |
|---|---|---|
| Example | NaCl | NH₄Cl |
| Scientific knowledge | Right | Right |
| Prior misconception | Right | Wrong |
| Expected ERP | P2↑ | N2↑ LPP↑ |
Existing behavioral research provides evidence indicating that self-explanation teaching intervention is effective in promoting conceptual change[20]. Meanwhile, neuroscience research reveals that the essence of conceptual change lies in the inhibition of misconceptions[10]. When performing tasks requiring suppression of misconception interference, inhibitory mechanisms are activated[21]. However, the neural activity profile directly linking self-explanation to misconception inhibition remains unestablished. This exploratory study aims to investigate whether intuitive conceptions about metallic compound composition persist in students' brains after acquiring scientific knowledge, and the impact of SES in rectifying misconceptions. Specifically, we hypothesized that:
(1) Incongruent stimuli compared to congruent stimuli will exist significant differences in behavioral results and ERP components (P2, N2, LPP).
(2) SSG compared to TIG will exist significant differences in behavioral results and related ERP components (P2, N2, LPP).
2. Review of the related literature
2.1. Students’ Misconceptions with Science Education
Research in science education has consistently highlighted the prevalence of students’ misconceptions, which are often resistant to traditional instruction. For instance, studies reveal that learners frequently hold intuitive yet scientifically inaccurate beliefs, such as the misconception that ionic compounds exclusively form between metal cations and non-metal anions. This is exemplified by students’ misinterpretation of compounds like NH₄Cl, where they erroneously attribute ionic bonding to metal-nonmetal interactions rather than recognizing polyatomic ions like NH₄⁺[22]. Such errors persist despite formal teaching, underscoring the robustness of intuitive frameworks.
Classical theoretical perspectives[23] posit that scientific concepts supplant prior conceptions through cognitive conflict resolution, emphasizing the necessity of conceptual replacement. Chi et al[14]. further proposed that conceptual change involves ontological recategorization. However, substantial evidence demonstrates the long-term coexistence of naive conceptions with scientific knowledge, requiring regulation through inhibitory mechanisms. The Prevalence Model accordingly confirms that multiple conceptions compete for cognitive dominance, with cognitive conflict optimally triggered post-instruction to facilitate conceptual transition[2].
Critically, inhibitory control mechanisms play a pivotal role in this coexistence framework. For example, when learners evaluate counterintuitive scientific statements, reaction times increase significantly due to interference from persistent misconceptions, reflecting the need for active inhibition[2,24]. Neural activity exhibits specific temporal dynamics during this process: activation of preconcepts occurs within 200-300 milliseconds, followed by conflict detection and resolution through inhibitory control, which suppresses intuitive responses to facilitate scientific conceptualization[5]. Key brain regions implicated include the dorsolateral prefrontal cortex (DLPFC), ventrolateral prefrontal cortex (VLPFC), and anterior cingulate cortex (ACC), which are more activated in experts during tasks requiring misconception suppression[10,21].
Nevertheless, pedagogical practices reveal a persistent conviction among educators that misconceptions can be fully eradicated through instruction. Traditional teaching methodologies predominantly focus on substituting correct concepts for erroneous ones, frequently failing to address misconception persistence. For instance, students taught ionic bonding through oversimplified models may retain core misunderstandings about electronegativity thresholds[25]. Consequently, effective interventions must incorporate strategies that systematically target the inhibition of intuitive responses. Evidence-based pedagogical approaches include: Refutational approaches that explicitly contrast scientific concepts with misconceptions through problem scenarios or refutational texts[21]; Inhibitory control training to suppress heuristic-based intuitive responses[5,9].
2.2. Self-Explanation Strategy
The SES refers to learners’ active generation of inferences extending beyond provided information to enhance comprehension of new knowledge[26]. Empirical evidence demonstrates a significant positive correlation between high-quality self-explanations and academic performance, constituting the “self-explanation effect” [27]. This metacognitive activity engages dual learning mechanisms: guiding learners to identify knowledge gaps while providing cognitive scaffolding and driving systematic reconstruction of mental models through integration processes[14,28]. When confronted with intentionally designed worked examples containing informational omissions, the self-explanation process stimulates learners to actively generate logical inferences that bridge cognitive gaps[14,29]. This constructive engagement compels continuous comparisons between existing cognitive frameworks and external information sources, triggering knowledge system reorganization upon detecting conceptual conflicts[30,31]. The strategy demonstrates cross-disciplinary applicability: In statics instruction, written self-explanation activities effectively support conceptual restructuring within classroom settings[32]; In computational science, self-explanation enhances students’ understanding and application of programming concepts[33]; In mathematics game design, self-explanation prompts linking gaming terminology to mathematical principles significantly improve learning outcomes[34].
In the context of overcoming deeply entrenched intuitive misconceptions, the theoretical significance of SES becomes particularly salient. Inhibitory control mechanisms are recognized as core cognitive processes for overcoming such misconceptions and achieving conceptual change, involving the suppression of intuitive erroneous responses or representations that conflict with scientific concepts[2,10,11]. Theoretically, self-explanation enhances this inhibitory process by facilitating deep processing and integration of conflicting information. Specifically, it primarily promotes conflict detection and monitoring: by requiring learners to reflect upon and elaborate learning materials, self-explanation improves metacognitive monitoring capacity, enabling more sensitive detection of inconsistencies between their understanding and presented scientific information or logical reasoning[14,35]—a prerequisite for triggering inhibitory control. Secondarily, it drives cognitive restructuring through explanation generation to bridge informational gaps or resolve conceptual conflicts, directly engaging in modifying or restructuring existing mental models[14,35]. This restructuring intrinsically attenuates activation strength of erroneous representations while modifying their connections with scientific concepts[20]. We thus posit that through active cognitive effort, self-explanation creates a more favorable internal cognitive environment for effective implementation of inhibitory control. While theoretical and behavioral studies confirm SES capacity to facilitate conceptual change, current research remains predominantly behaviorally oriented, lacking mechanistic insights into the neural substrates underlying these cognitive transformations.
2.3. Event-Related Potential Technology
Event-Related Potential technology refers to a non-invasive electrophysiological method used to measure brain activity associated with specific sensory, cognitive, or motor events. ERPs are derived from the electroencephalogram (EEG) by averaging neural responses time-locked to repeated occurrences of an event, allowing researchers to isolate consistent neural activity from background noise[16]. When individuals perform cognitive tasks, different types of stimuli can affect the activation patterns of relevant neural networks, leading to characteristic changes in ERP waveforms. The P200 component serves as an indicator of expectancy processes, reflecting comparisons between perception and existing memory. A match between these elements results in a positive P200 amplitude[17]. A study investigating visual learning revealed that the P200 component evoked by familiar words exhibited a more positive response than that evoked by novel words[36]. The N200, a negative component emerging at 200–350 ms, is linked to conflict monitoring and error detection[18]. Research conducted in go/no-go experiments has shown that the N200 component is more pronounced in response to no-go stimuli[37]. Stimulus conditions that induce semantic conflicts result in conflict effects, leading to larger N200 components[38]. The LPP component measures cognitive processes related to conscious error recognition, conflict resolution, and response selection[39]. Retrieval of scientific knowledge, detection of conflicts, and suppression of misconceptions lead to increased LPP amplitudes during problem-solving[40]. In summary, analyzing the different EEG signal characteristics exhibited by subjects under various stimulus conditions can provide insights into their cognitive processes. Therefore, analyzing distinct ERP components (e.g., P2, N2, LPP) under varying stimulus conditions provides a powerful non-invasive method to elucidate the temporal dynamics of cognitive processes, including conflict detection, inhibitory control, central to the aims of this study.
3. Method
3.1. Participants
A total of 23 first-year high school students from City X (15 males, 8 females), aged 16, were recruited for this study. All participants were physically and mentally healthy, with no history of brain injury, substance abuse, or psychiatric disorders, were right-handed, and had normal vision or corrected vision. Each participant signed an informed consent form before the experiment and received appropriate compensation. The participants were randomly and equally assigned to two groups: the SSG and the TIG. In the SSG, participants were asked to provide self-explanations after completing each instructional video on Chapter 4, Section 3 (“The Structure of Substances—The Periodic Law”) of the People’s Education Press Chemistry Book Ⅰ, based on prompts provided. Participants in the TIG read all provided concepts after finishing each video. Subsequently, data were screened to exclude non-compliant data, resulting in a final sample of 15 participants. Eight participants were excluded due to excessive EEG artifacts or failure to follow self-explanation protocols.
3.2. Stimuli
This investigation focused on high school students’ cognitive development regarding chemical bonding, specifically targeting the prevalent misconception that “ionic compounds exclusively consist of metal and nonmetal elements.” We developed a set of contrasting stimuli: Congruent stimuli: Presented 19 prototypical ionic compounds aligning with students’ intuitive conceptions (e.g., NaCl, comprising metal cation Na⁺ and nonmetal anion Cl⁻); Incongruent stimuli: Featured 3 non-canonical ionic compounds contradicting intuitive beliefs (e.g., NH₄Cl, containing nonmetal polyatomic cation NH₄⁺ paired with nonmetal anion Cl⁻). Congruent and incongruent stimuli were randomized. To prevent habitual responding, we interspersed 21 irrelevant stimuli comprising 17 covalent compounds (e.g., CO₂) and 4 neutral compounds (neither ionic nor covalent).
3.3. Experimental procedure
Upon arrival at the EEG laboratory, participants were required to have washed and thoroughly dried their hair and scalp beforehand. They were then familiarized with the laboratory environment and asked to complete a basic information questionnaire. Next, the EEG cap was introduced to the participants, along with an explanation of the safety of the experimental equipment. The EEG cap was then fitted to the participants, ensuring that it met all the requirements for accurate ERP data collection. Before the experiment began, the procedure was explained to the participants, with instructions to relax their head and facial muscles as much as possible and to minimize blinking. The specific tasks of the experiment were also clarified, emphasizing the need for quick responses during the task. Participants were seated comfortably in front of a computer screen, ensuring their eyes were level with the screen and maintaining approximately 60 centimeters. Following this, participants were guided through the operation of the response keys and the entire experimental process to ensure they were fully prepared and understood the requirements (Figure 1). Assistance was provided if needed until the participant was completely familiar with the procedure.
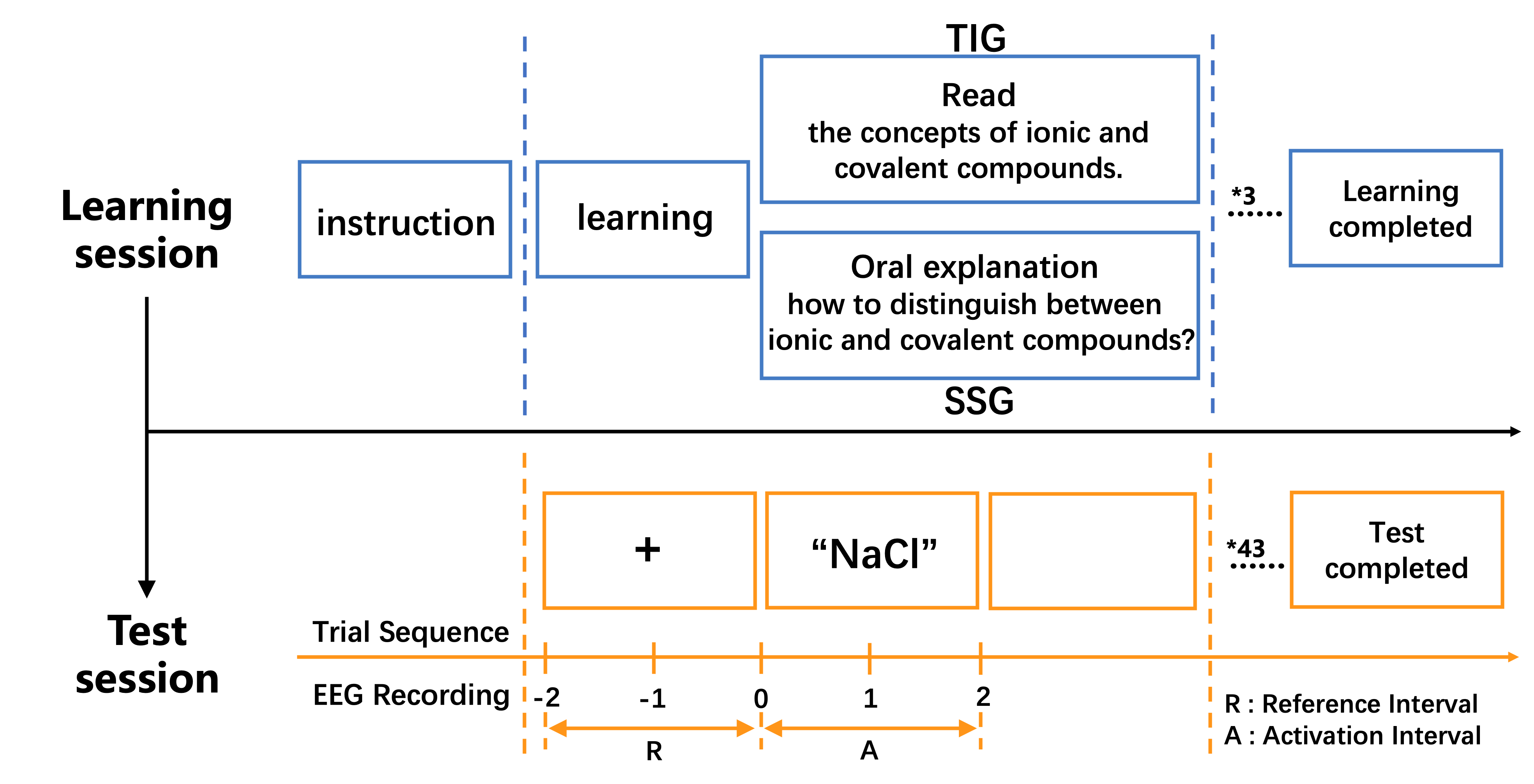
Figure 1. A flow chart of experimental research
The experiment began with a 2000 ms presentation of a fixation cross at the center of the screen, followed by video-based learning materials. After each video segment, SSG received self-explanation prompts, while the TIG reviewed conceptual summaries corresponding to the video content. Upon completing the first video session, participants pressed a key to advance to the next video, repeating this process for three sequential learning modules. In the subsequent stimulus evaluation phase, a fixation cross reappeared for 2000 ms, after which stimulus materials were displayed for 2000 ms. Participants judged the validity of each stimulus by pressing the “T” key for correct classifications or the “F” key for incorrect classifications. Trials without responses (0.8% of total data) were excluded as invalid. This procedure continued until all 43 stimulus sets were presented, concluding the experimental session. Response mappings and trial sequences were counterbalanced across participants to control for order effects.
3.4. Behavioral data analysis
First, data from the TIG and SSG were collected using E-Prime 2.0 software. The data were then merged using E-Merge and E-Data Aid, and subsequently imported into Excel 2021 for preprocessing. During this stage, accuracy and reaction time were calculated (with incorrect responses filtered during reaction time computation). Data from the practice phase and incorrect responses were removed. The final processed data were imported into SPSS 27.0 for statistical analysis.
3.5. EEG recording and data analysis
The Neuracle EEG system was used to collect ERP data during the experimental process, and the resulting data were converted into a format that could be processed using MATLAB. The data were then analyzed using the EEGLAB toolbox.
Based on previous relevant literature and observations of the grand average ERP waveforms and topographical maps from this study, nine electrodes were selected for focused analysis (frontal region: F3, F4, Fz; parietal region: P3, Pz, P4; occipital region: PO7, Oz, PO8). After extracting the relevant ERP data, the data were imported into SPSS 27.0 for statistical analysis. A 3 (lateral regions: F3, Fz, F4) × 2 (stimulus conditions: I, CI) repeated measures ANOVA was conducted on the P2 and N2 components for both the SSG and TIG. For the LPP component, a 3 (P3, Pz, P4) × 2 (I, CI) repeated measures ANOVA. The p-values for all ANOVAs were adjusted using the Greenhouse-Geisser correction, and Bonferroni correction was applied for post hoc multiple comparisons.
4. Results
4.1. Behavioral results
The mean response accuracy and reaction times across stimulus conditions for all 15 participants were analyzed (Table 2). Accuracy distributions for both groups under each condition violated normality assumptions (Shapiro-Wilk tests: p<.05), warranting non-parametric statistical analyses. Wilcoxon signed-rank tests revealed no significant differences between the TIG and SSG in either C (z=-0.169, p=.86) or IC (z=-0.106, p=.92) conditions. Kruskal-Wallis tests further confirmed no group-level accuracy differences for either stimulus type (C: χ² (1) = 0.372, p=.54; IC: χ² (1)=0.066, p=.80).
Table 2. Behavioral Performance and ERP Descriptive Results for Each Condition
| Group | Condition | Response time | Accuracy rate | Component | Amplitude |
|---|---|---|---|---|---|
| TIG | C | 862.15±157.97 | 84.87±18.33 | P200 | 4.39±3.23 |
| N200 | -0.56±2.74 | ||||
| LPP | 0.81±3.35 | ||||
| IC | 1014.31±159.81 | 75.00±34.50 | P200 | 0.37±3.69 | |
| N200 | -3.61±4.28 | ||||
| LPP | 1.57±4.08 | ||||
| SSG | C | 886.40±139.33 | 92.48±13.19 | P200 | 2.08±2.35 |
| N200 | -3.56±3.45 | ||||
| LPP | 0.07±1.28 | ||||
| IC | 914.10±192.52 | 80.95±26.23 | P200 | 0.66±2.40 | |
| N200 | -2.92±1.57 | ||||
| LPP | 1.61±4.87 |
Shapiro-Wilk tests confirmed normal distributions for reaction times across groups and stimulus types (p-values ranging from .44 to .83), satisfying parametric analysis assumptions. A mixed-design ANOVA revealed a marginally significant main effect of stimulus condition on reaction time (F (1,13)=3.307, p=.092), with no other significant main effects or interactions (all p>.5). Post hoc simple effects analysis demonstrated that the TIG exhibited significantly longer reaction times for IC compared to C (p<.05), whereas the SSG showed no significant reaction time differences between stimulus conditions.
4.2. Neurophysiological Results
Grand average ERP waveforms revealed distinct component activations across experimental conditions. The P2 component exhibited a positive peak around 200 ms post-stimulus, with maximal amplitudes observed in frontal regions. Mean P2 amplitudes were quantified within the 150–240 ms time window. The N2 component reached its negative peak at approximately 290 ms, localized to frontal electrodes, with mean amplitudes analyzed between 250–350 ms. The late positive potential (LPP) demonstrated maximal amplitude in parietal regions during the 700–900 ms window.
Figure 2 illustrates the TIG’s midline ERP waveforms and topographic maps of P2, N2, and LPP for C and IC trials. For the P2 component, a repeated-measures ANOVA revealed a significant main effect of stimulus type (F(1,13) =25.352, p<.001, η²=0.547), with C stimuli eliciting significantly larger amplitudes than IC stimuli. No significant main effects of electrode location or stimulus-electrode interactions were observed (all p>.5). The N2 analysis showed a significant main effect of stimulus type (F(1,13)=6.075, p=.022, η²=0.224), with IC stimuli generating larger N2 amplitudes than C stimuli, without other significant effects (all p>.5). For the LPP, significant main effects of stimulus type (F(1,13)=9.528, p<.01, η²=0.312) and electrode location (F(2,26)=3.770, p<.05, η²=0.264) emerged. Pairwise comparisons indicated larger amplitudes at Pz compared to P3 (p<.05), with no other inter-electrode differences. A significant stimulus-electrode interaction (F(2,26)=5.025, p<.05, η²=0.324) was observed, where simple effects analysis revealed greater IC-induced LPP amplitudes at P3 compared to C stimuli.
Figure 3 displays the SSG’s midline ERP waveforms and component topographies. For the P2, a two-way repeated-measures ANOVA detected no significant main effects or interactions (all p>.5). Similarly, N2 analysis yielded no statistically significant outcomes (all p>.5). The LPP analysis demonstrated a significant main effect of electrode location (F(2,26)=3.883, p<.05, η²=0.301), with Pz amplitudes exceeding those at P3 and P4 (p<.05). No other significant effects were observed (all p>.5).
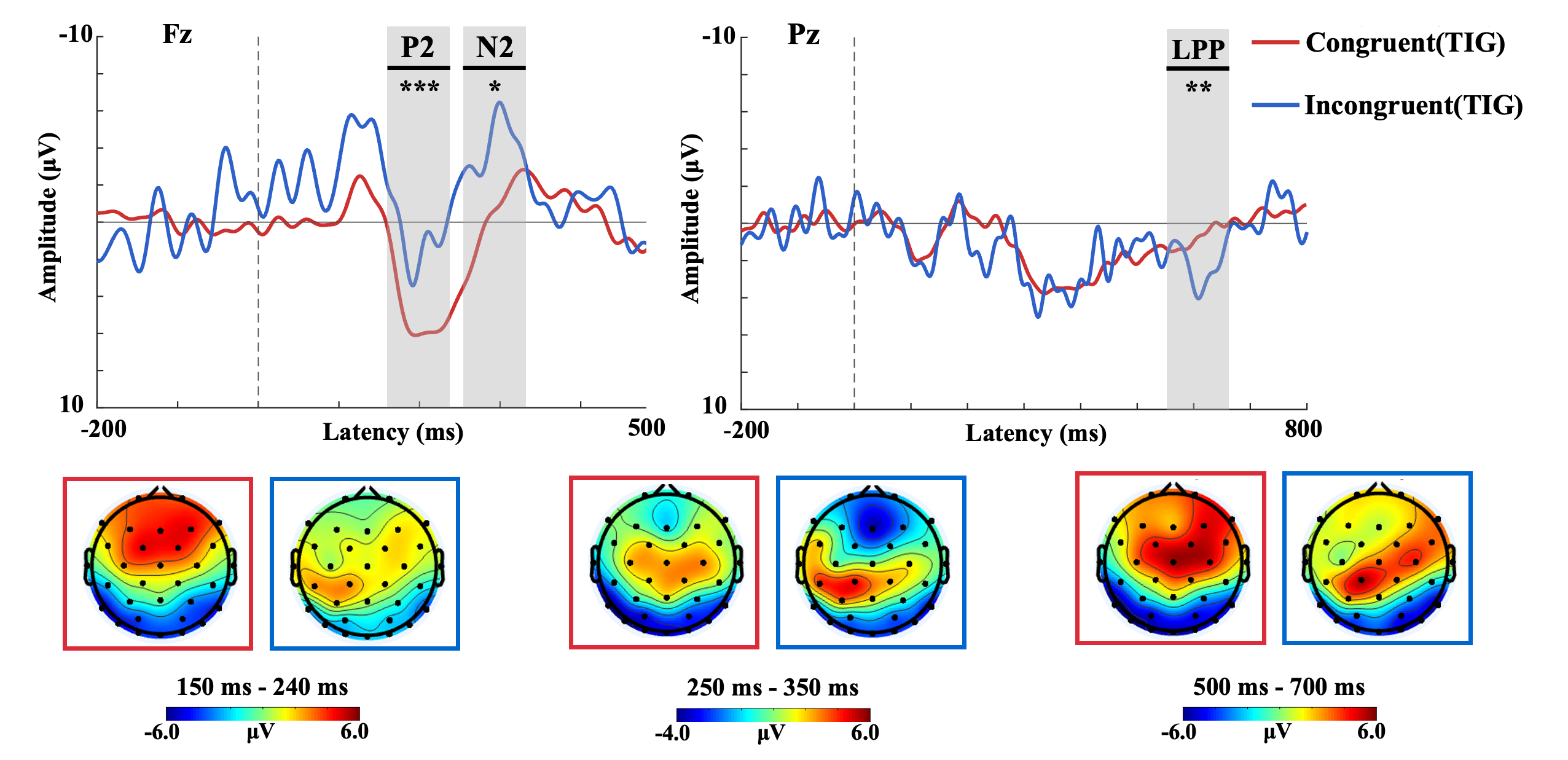
Figure 2. The ERP waveforms of the TIG in midline electrodes and topographical maps of P2, N2, and LPP in congruent and incongruent trials.
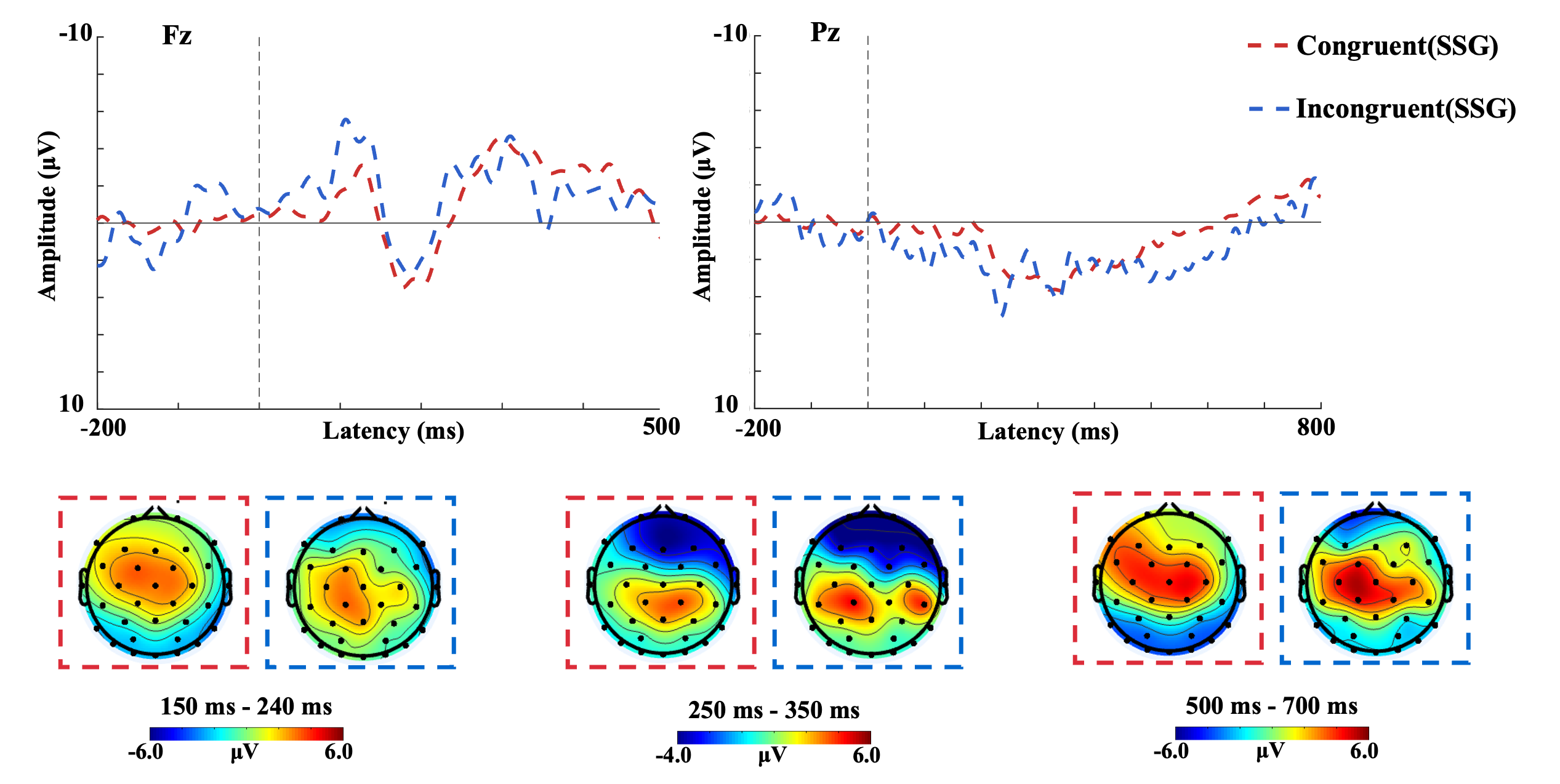
Figure 3. The ERP waveforms of the SSG in midline electrodes and topographical maps of P2, N2, and LPP in congruent and incongruent stimuli.
Comparative analyses of P2 component differences between the TIG and SSG across stimulus conditions were conducted using repeated-measures ANOVA on P200 amplitudes at distinct electrode sites (Figure 4). For C stimuli, a marginally significant main effect of group was observed (F(1,13) = 4.62, p=.051, η²=0.262), with no significant main effects of electrode location or group-electrode interactions (all p>.5). In contrast, analyses of IC stimuli revealed no significant main effects of group or electrode (all p>.5), but a significant group-electrode interaction emerged in multivariate testing (F(2,12) = 4.453, p<.05, η²=0.426). Subsequent simple effects analysis identified a significant linear trend difference at electrode F4 between groups (F(1,13)=9.347, p=.009, η²=0.418).
For N2 component comparisons, repeated-measures ANOVA of N2 amplitudes demonstrated distinct patterns (Figure 4). Under C stimulation, significant main effects of group (F(1,13)=7.686, p<.05, η²=0.372) and electrode location (F(2,26)=4.944, p<.05, η²=0.276) were observed. Specifically, the SSG exhibited significantly more negative N2 amplitudes than the TIG, with reduced amplitudes at Fz compared to F4. No significant interactions were detected. For IC stimuli, no significant main effects or interactions emerged (all p>.5).
LPP component analyses revealed significant electrode effects for both stimulus types (Figure 4). Under C stimulation, a main effect of electrode location was significant (F(2,26)=7.347, p<.01, η²=0.361), with pairwise comparisons showing larger amplitudes at Pz than P3 and P4. No group effects or interactions were observed (all p>.5). For IC stimuli, significant electrode effects persisted (F(2,26)=7.382, p<.01, η²=0.362), with Pz and P3 amplitudes exceeding P4. No other significant effects were detected (all p>.5).
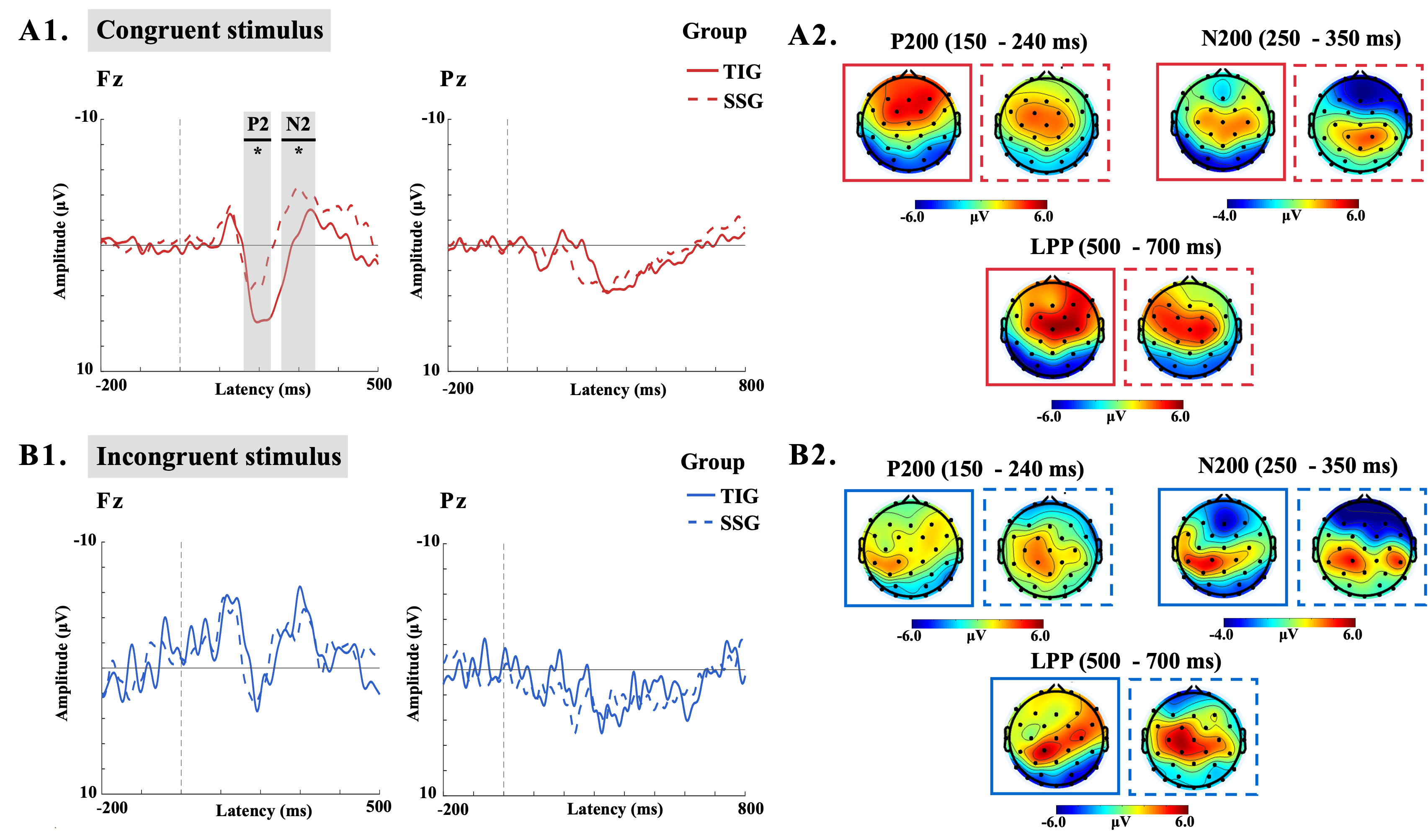
Figure 4. (a) The ERP waveforms for the TIG and SSG groups across both Congruent Trials (A1) and Incongruent Trials (B1) are displayed at Fz and Pz sites; (b) Topographical maps of the P2, N2, and LPP components for the TIG and SSG across both Congruent Trials (A2) and Incongruent Trials (B2).
5. Discussion
This study aimed to investigate the persistence of students' intuitive misconception that “ionic compounds exclusively consist of metal cations and nonmetal anions” following instruction, specifically examining its manifestation through characteristic EEG inhibitory control patterns during atypical ionic compound recognition. Behavioral and EEG analyses appeared to confirm this hypothesis. Furthermore, we investigated the impact of the SES on correcting such misconceptions. Collectively, the behavioral and electrophysiological data suggest that self-explanation may weaken the activation of intuitive concepts during early scientific reasoning while enhancing conflict monitoring and inhibitory control.
In the TIG, behavioral data revealed prolonged reaction times for IC stimuli compared to C stimuli. This aligns with prior evidence linking inhibitory demands to extended RTs[15], supporting our prediction that resolving IC stimuli requires suppressing intuitive conceptions of ionic compound composition. However, the absence of accuracy differences between conditions may reflect task simplicity rather than equivalent conceptual processing.
ERP findings in the TIG further corroborated the conceptual coexistence theory. First, enhanced P2 amplitudes for C stimuli likely index higher congruence with pre-existing intuitive schemas. As an anticipatory marker, the P2 component reflects implicit comparisons between sensory input and memory-based expectations[17]. The larger P2 amplitudes for prototypical ionic compounds (C stimuli) suggest heightened perceptual alignment with intuitive misconceptions during early-stage scientific processing. Second, increased N2 amplitudes for IC stimuli implicate anterior cingulate cortex (ACC)-mediated conflict monitoring[41], indicating automatic detection of conceptual mismatches. Finally, amplified LPP amplitudes for IC stimuli, localized to parietal regions, signify conscious conflict resolution and effortful retrieval of scientific knowledge to override intuitive responses[19]. Collectively, these neural signatures confirm the persistence of intuitive conceptions post-instruction, necessitating sustained inhibition during conceptual evaluation.
The SSG exhibited distinct neural profiles, with no significant ERP differences between C and IC stimuli across components (P2/N2/LPP), paralleling the absence of behavioral performance distinctions. The lack of reaction time prolongation for IC stimuli in the SSG may reflect optimized inhibitory efficiency.
To further elucidate the intervention effects and neural mechanisms of the SES on misconception mitigation, we conducted between-group comparisons of ERP components under distinct stimulus conditions. Critically, under C stimulus conditions, SES significantly modulated early ERP responses in the SSG compared to the TIG. The reduced P2 amplitude observed in the SSG suggests that SES weakened the automatic activation and dominance of intuitive conceptual representations. This diminished P2, a marker of stimulus-concept expectation matching, indicates that SES reduces the neural salience and default reliance on intuitive knowledge when processing scientifically congruent information.
Furthermore, the SSG exhibited enhanced N2 amplitudes in response to C stimuli. This finding is particularly significant as C stimuli are typically not expected to elicit conflict. The heightened N2, associated with conflict monitoring and cognitive control processes[18], demonstrates that SES training fundamentally alters cognitive processing. It appears to induce a state of heightened vigilance or conflict detection even when presented with information that superficially aligns with prior (intuitive) expectations. This enhanced early conflict monitoring potentially reflects a key mechanism by which SES fosters conceptual change – by making learners more sensitive to potential discrepancies, even in seemingly straightforward situations.
The combined effect of reduced P2 and enhanced N2 amplitudes under C conditions provides direct neural evidence that SES training modulates early stages of scientific thinking by attenuating the automatic activation strength of intuitive conceptions and enhancing conflict monitoring mechanisms. This shift in neural processing dynamics – weaker initial conceptual matching but stronger early conflict detection – aligns with behavioral observations of SES promoting deeper processing and reducing reliance on potentially flawed intuitive models[14,42]. It suggests that SES fosters a more critical and controlled approach to information processing, even for concepts that appear intuitively correct.
In contrast, for IC stimuli, no significant differences in P2, N2, or LPP amplitudes were found between the TIG and SSG. This indicates that SES training, as implemented here, exerted minimal influence on the neural correlates of conflict resolution specifically when suppressing strong, pre-existing misconceptions during task performance. The resolution of this overt conflict may rely on mechanisms not differentially modulated by our SES intervention compared to traditional instruction.
6. Conclusions
This study investigated the persistence of intuitive conceptions regarding ionic compound composition post-scientific instruction and the neurocognitive mechanisms of SES in misconception correction by integrating behavioral data and ERP technology. The findings validate the conceptual coexistence theory, demonstrating that intuitive misconceptions persist in cognitive architecture after scientific knowledge acquisition, engaging inhibitory mechanisms during scientific reasoning. Crucially, our results reveal that the SES may attenuate early-stage intuitive concept activation while amplifying cognitive control demands, thereby facilitating proactive inhibition of entrenched misconceptions.
These insights carry significant implications for educators and science pedagogy. The “learning from errors” paradigm necessitates systematic analysis of common misconceptions to identify their origins, assess conceptual understanding, and address structural knowledge gaps. Given the persistent interference of intuitive conceptions, educators should design opportunities for knowledge application that strengthen students’ capacity to detect and suppress erroneous beliefs. We propose integrating self-explanation prompts with inhibitory training—specifically embedding explanatory cues at conceptual conflict junctures—to optimize inhibitory control over misconceptions.
Theoretical contributions are twofold: First, neural evidence substantiates the enduring nature of domain-specific intuitive misconceptions, providing cross-disciplinary validation for the conceptual coexistence framework. Second, the study elucidates mechanistic pathways through which self-explanation fosters conceptual change. Practically, these findings empirically support self-explanation-based pedagogical interventions, emphasizing the importance of inhibitory training in science education.
Limitations include a moderate sample size and constrained task complexity. Future research should extend these investigations to diverse disciplinary contexts, employ longitudinal designs to assess intervention sustainability, and incorporate fMRI to map functional connectivity changes in inhibition-related neural networks.
Conflicts of Interest:
The authors declare no conflict of interest.
Funding:
This work was supported by the National Natural Science Foundation of China (NSFC) under grant number [72004166].
Author contributions:
Jiekai Li: First author, Formal analysis, Software, Visualization, Writing Original Draft. Yu Teng, Yinghao Jin and Mengnan Zhang: Second, Third and Forth author, Investigation, Revising Original Draft. Jianqiang Ye: Fifth author and Corresponding author, Conceptualization, Methodology, Writing Review.
References
-
J. Nussbaum, S. Novick. "Alternative Frameworks, Conceptual Conflict and Accommodation: Toward a Principled Teaching Strategy." Appl. Phys. Lett. 1982, 11, 3, 183-200.
-
P. Potvin, S. Masson, S. Lafortune, G. Cyr. "PERSISTENCE OF THE INTUITIVE CONCEPTION THAT HEAVIER OBJECTS SINK MORE: A REACTION TIME STUDY WITH DIFFERENT LEVELS OF INTERFERENCE." Int J of Sci and Math Educ 2015, 13, 1, 21-43.
-
M. C. Periago, X. Bohigas. "A study of second-year engineering students' alternative conceptions about electric potential, current intensity and Ohm's law." Appl. Phys. Lett. 2005, 30, 1, 71-80.
-
A. Shtulman, J. Valcarcel. "Scientific knowledge suppresses but does not supplant earlier intuitions." Cognition 2012, 124, 2, 209-15.
-
Y. Zhu, L. Zhang, Y. Leng, R. Pang, X. Wang. "Event-Related Potential Evidence for Persistence of an Intuitive Misconception About Electricity." Appl. Phys. Lett. 2019, 13, 2, 80-91.
-
Y. Skelling-Desmeules, L.-M. Brault Foisy, P. Potvin, H. G. Lapierre, E. Ahr, P.-M. Léger, S. Masson, P. Charland. "Persistence of the 'Moving Things Are Alive' Heuristic into Adulthood: Evidence from EEG." LSE 2021, 20, 3, ar45.
-
R. F. Goldberg, S. L. Thompson-Schill. "Developmental 'Roots' in Mature Biological Knowledge." Psychol Sci 2009, 20, 4, 480-87.
-
J. Evans, K. Stanovich. "Dual-Process Theories of Higher Cognition." Perspectives on Psychological Science 2013, 8, 223-41.
-
O. Houdé, G. Borst. "Measuring inhibitory control in children and adults: brain imaging and mental chronometry." Front. Psychol. 2014, 5.
-
S. Masson, P. Potvin, M. Riopel, L.-M. B. Foisy. "Differences in Brain Activation Between Novices and Experts in Science During a Task Involving a Common Misconception in Electricity." Appl. Phys. Lett. 2014, 8, 1, 44-55.
-
L.-M. Brault Foisy, P. Potvin, M. Riopel, S. Masson. "Is inhibition involved in overcoming a common physics misconception in mechanics?" Trends in Neuroscience and Education 2015, 4, 1, 26-36.
-
G. Allaire-Duquette, M. Bélanger, R. H. Grabner, K. Koschutnig, S. Masson. "Individual differences in science competence among students are associated with ventrolateral prefrontal cortex activation." Appl. Phys. Lett. 2019, 97, 9, 1163-78.
-
M. Bélanger, P. Potvin, S. Horstis, A. Shtulman, E. F. Mortimer. "Multidisciplinary perspectives on representational pluralism in human cognition." Appl. Phys. Lett. 2023.
-
M. T. Chi. "Self-explaining expository texts: The dual processes of generating inferences and repairing mental models." Appl. Phys. Lett. 2013, 161-238.
-
M. M. Botvinick, T. S. Braver, D. M. Barch, C. S. Carter, J. D. Cohen. "Conflict monitoring and cognitive control." Appl. Phys. Lett. 2001, 108, 3, 624.
-
S. J. Luck. "An introduction to the event-related potential technique." Appl. Phys. Lett. 2014.
-
K. M. Evans, K. D. Federmeier. "The memory that's right and the memory that's left: Event-related potentials reveal hemispheric asymmetries in the encoding and retention of verbal information." Appl. Phys. Lett. 2007, 45, 8, 1777-90.
-
J. R. Folstein, C. Van Petten. "Influence of cognitive control and mismatch on the N2 component of the ERP: A review." Psychophysiology 2008, 45, 1, 152-70.
-
E. Coderre, K. Conklin, W. J. van Heuven. "Electrophysiological measures of conflict detection and resolution in the Stroop task." Appl. Phys. Lett. 2011, 1413, 51-59.
-
K. Bisra, Q. Liu, J. C. Nesbit, F. Salimi, P. H. Winne. "Inducing Self-Explanation: a Meta-Analysis." Educ Psychol Rev 2018, 30, 3, 703-25.
-
L. Mason, S. Zaccoletti. "Inhibition and Conceptual Learning in Science: a Review of Studies." Educ Psychol Rev 2021, 33, 1, 181-212.
-
A. K. Prodjosantoso, A. M. Hertina, Irwanto. "The Misconception Diagnosis on Ionic and Covalent Bonds Concepts with Three Tier Diagnostic Test." Appl. Phys. Lett. 2019, 12, 1, 1477-88.
-
G. J. Posner, K. A. Strike, P. W. Hewson, W. A. Gertzog. "Accommodation of a scientific conception: Toward a theory of conceptual change." Appl. Phys. Lett. 1982, 66, 2, 211-27.
-
R. Babai, A. Amsterdamer. "The Persistence of Solid and Liquid Naive Conceptions: A Reaction Time Study." J Sci Educ Technol 2008, 17, 6, 553-59.
-
R. Vladušić, R. B. Bucat, M. Ožić. "Understanding ionic bonding-a scan across the Croatian education system." Appl. Phys. Lett. 2016, 17, 4, 685-99.
-
M. T. Chi, M. Bassok, M. W. Lewis, P. Reimann, R. Glaser. "Self-explanations: How students study and use examples in learning to solve problems." Appl. Phys. Lett. 1989, 13, 2, 145-82.
-
M. Roy, M. T. H. Chi. "The Self-explanation Principle." Appl. Phys. Lett. .
-
J. R. Morrison, L. Bol, S. M. Ross, G. S. Watson. "Paraphrasing and prediction with self-explanation as generative strategies for learning science principles in a simulation." Education Tech Research Dev 2015, 63, 6, 861-82.
-
R. G. Hausmann, K. VanLehn. "The effect of self-explaining on robust learning." Appl. Phys. Lett. 2010, 20, 4, 303-32.
-
L. Fiorella, R. E. Mayer. "Learning as a generative activity: Eight learning strategies that promote understanding." Appl. Phys. Lett. 2015.
-
A. Renkl, R. Stark, H. Gruber, H. Mandl. "Learning from worked-out examples: The effects of example variability and elicited self-explanations." Appl. Phys. Lett. 1998, 23, 1, 90-108.
-
J. L. De La Hoz, C. Vieira, C. Arteta. "Self‐explanation activities in statics: A knowledge‐building activity to promote conceptual change." J of Engineering Edu 2023, 112, 3, 741-68.
-
C. Vieira, A. J. Magana, M. L. Falk, R. E. Garcia. "Writing In-Code Comments to Self-Explain in Computational Science and Engineering Education." ACM Trans. Comput. Educ. 2017, 17, 4, 1-21.
-
H. F. O'Neil, G. K. Chung, D. Kerr, T. P. Vendlinski, R. E. Buschang, R. E. Mayer. "Adding self-explanation prompts to an educational computer game." Appl. Phys. Lett. 2014, 30, 23-28.
-
D. S. McNamara, J. P. Magliano. "Self-explanation and metacognition: The dynamics of reading." Appl. Phys. Lett. 2009, 60-81.
-
T. Curran, J. Dien. "Differentiating amodal familiarity from modality‐specific memory processes: An ERP study." Psychophysiology 2003, 40, 6, 979-88.
-
S. Nieuwenhuis, N. Yeung, W. Van Den Wildenberg, K. R. Ridderinkhof. "Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency." Cognitive, Affective, & Behavioral Neuroscience 2003, 3, 1, 17-26.
-
P. Kanske, S. A. Kotz. "Modulation of early conflict processing: N200 responses to emotional words in a flanker task." Appl. Phys. Lett. 2010, 48, 12, 3661-64.
-
K. A. Paller, J. L. Voss, S. G. Boehm. "Validating neural correlates of familiarity." Appl. Phys. Lett. 2007, 11, 6, 243-50.
-
H. Yin, D. Li, J.- Yang, W. Li, J. Qiu, Y. Chen. "Neural bases of unconscious error detection in a Chinese anagram solution task: Evidence from ERP study." Appl. Phys. Lett. 2016, 11, 5, e0154379.
-
V. van Veen, C. S. Carter. "The timing of action-monitoring processes in the anterior cingulate cortex." Appl. Phys. Lett. 2002, 14, 4, 593-602.
-
V. A. W. M. M. Aleven, K. R. Koedinger. "An effective metacognitive strategy: learning by doing and explaining with a computer‐based Cognitive Tutor." Cognitive Science 2002, 26, 2, 147-79.